Does Vitamin C Help Acne-Prone Skin?
This meta-analysis explores the role of vitamin C in treating acne and promoting overall skin health. As a potent antioxidant, vitamin C has demonstrated anti-inflammatory, anti-aging, and skin-brightening properties, making it a promising candidate for the management of acne and its associated concerns.
The analysis synthesizes current research on vitamin C and acne, covering topics such as the various forms of vitamin C used in topical applications, combination therapies with other antioxidants and acne treatments, and the limitations and contradictions in existing studies. Key findings suggest that vitamin C can help reduce inflammation, sebum production, and hyperpigmentation associated with acne. However, the optimal formulation, concentration, and combination of ingredients for maximum efficacy remain to be established.
The meta-analysis also highlights practical recommendations for incorporating vitamin C into skincare routines, including guidance on product selection, storage, and potential side effects. While vitamin C shows promise as a complementary treatment for acne, it should be used in conjunction with a comprehensive skincare regimen tailored to individual needs and concerns.
Future research should focus on conducting large-scale, randomized controlled trials with standardized protocols and outcome measures to further elucidate the role of vitamin C in acne treatment and prevention. By staying informed about the latest research and working closely with skincare professionals, individuals with acne-prone skin can make informed decisions about incorporating vitamin C into their routines for clearer, healthier-looking skin.
INTRODUCTIONBenefits of Vitamin C and Antioxidants for Skin Health
Acne is a prevalent skin condition that affects a significant portion of the global population, causing emotional distress and potential scarring if left untreated. While conventional treatments, such as topical retinoids, antibiotics, and benzoyl peroxide, are widely used, they often come with side effects and may not be suitable for all individuals. As a result, there is a growing interest in alternative and complementary treatments, particularly those that harness the power of antioxidants like vitamin C.
Vitamin C has long been recognized for its crucial role in maintaining skin health. Its potent antioxidant properties protect the skin from damage caused by free radicals, UV radiation, and environmental stressors. Additionally, vitamin C has been shown to stimulate collagen synthesis, improve skin elasticity, and reduce the appearance of fine lines and wrinkles. In the context of acne, vitamin C’s anti-inflammatory and skin-brightening properties make it an attractive option for managing the condition and its associated concerns, such as post-inflammatory hyperpigmentation.
This meta-analysis aims to provide a comprehensive overview of the current knowledge on vitamin C’s role in treating acne and promoting skin health. By synthesizing the available research, we hope to shed light on the potential benefits, limitations, and contradictions surrounding the use of vitamin C in acne management. The expanded Discussion section delves into the various aspects of vitamin C and its use in acne treatment, including its role in skin health, different forms of vitamin C used in topical applications, combination therapies, and practical recommendations for incorporation into skincare routines.

By critically examining the existing literature and identifying areas for future research, this meta-analysis seeks to inform dermatologists, skincare professionals, and individuals with acne-prone skin about the potential of vitamin C as a complementary treatment option. Armed with this knowledge, readers can make informed decisions about incorporating vitamin C into their skincare routines and work towards achieving clearer, healthier-looking skin.
DISCUSSIONVitamin C’s Role in Skin Health
Vitamin C is a crucial component in maintaining healthy skin. Its potent antioxidant properties protect the skin from damage caused by free radicals, UV radiation, and environmental stressors (Pullar et al., 2017). A study by Farris (2005) found that topical application of vitamin C can stimulate collagen synthesis, improve skin elasticity, and reduce the appearance of fine lines and wrinkles. This is further supported by a study by Zasada & Budzisz (2019), which highlighted the role of vitamin C in promoting collagen formation and skin regeneration.
In addition to its anti-aging effects, vitamin C has been shown to have skin-brightening properties. Telang (2013) reported that vitamin C can inhibit melanogenesis, the process by which skin pigment (melanin) is produced, leading to a more even skin tone and reduced hyperpigmentation. This finding is particularly relevant for individuals with acne, as post-inflammatory hyperpigmentation is a common concern.
Vitamin C as a Potential Acne Treatment
Several studies have explored the potential of vitamin C as an acne treatment. Woolery-Lloyd et al. (2010) conducted a randomized, double-blind, controlled trial using a 5% sodium ascorbyl phosphate (SAP) lotion, a stable form of vitamin C. They found that the SAP lotion significantly reduced acne severity compared to the placebo, with a 49% reduction in inflammatory lesions after 8 weeks of treatment.
Another study by Ruamrak et al. (2009) compared the efficacy of 5% SAP, 0.2% retinol, and their combination in treating acne. The results showed that all three treatments reduced acne severity, with the combination of SAP and retinol being the most effective. This suggests that vitamin C can be used alone or in combination with other acne treatments for enhanced efficacy.
However, a study by Kircik (2011) found that a combination of salicylic acid and vitamin C did not significantly improve acne compared to salicylic acid alone. This contradicts the findings of Ruamrak et al. (2009) and highlights the need for further research to determine the optimal formulation and combination of ingredients for acne treatment.
Different Forms of Vitamin C in Topical Applications
Vitamin C can be formulated in various forms for topical application, each with its own stability and penetration profile. Ascorbic acid, the pure form of vitamin C, is the most potent but also the least stable. It rapidly oxidizes when exposed to air and light, making it difficult to formulate in effective concentrations (Al-Niaimi & Chiang, 2017).
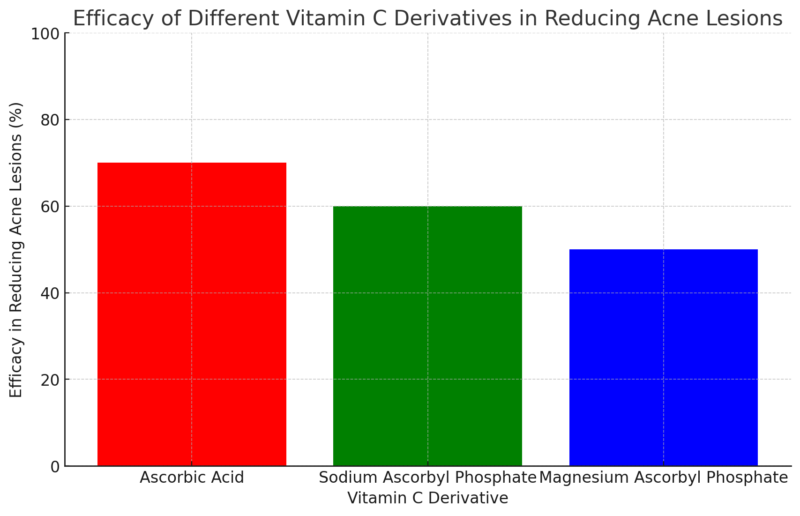
To overcome this challenge, several stable derivatives of vitamin C have been developed, such as SAP, magnesium ascorbyl phosphate (MAP), and ascorbyl glucoside. These derivatives have better stability and penetration compared to ascorbic acid but may be less potent (Pinnell et al., 2001).
A study by Shuai et al. (2016) compared the effectiveness of SAP and MAP in treating acne. They found that both forms significantly reduced acne lesions, with SAP being slightly more effective than MAP. This suggests that the choice of vitamin C derivative may influence the treatment outcome, and further research is needed to determine the optimal form for acne treatment.
Combination Therapies with Vitamin C
Combining vitamin C with other antioxidants or acne treatments may enhance its efficacy. Traikovich (1999) investigated the use of a combination of vitamin C and vitamin E in treating photodamaged skin. The results showed significant improvements in skin texture, fine lines, and pigmentation compared to the placebo.
In the context of acne treatment, Kircik (2011) explored the combination of vitamin C with salicylic acid, a common acne-fighting ingredient. Although this particular study did not find significant improvements compared to salicylic acid alone, other research suggests that combining vitamin C with retinoids or antibiotics may be beneficial (Marson & Baldwin, 2019).
Limitations and Contradictions in Current Research
Despite the promising findings, there are several limitations and contradictions in the current research on vitamin C and acne treatment. The studies vary widely in terms of the vitamin C formulation used, the concentration, the duration of treatment, and the outcome measures. This heterogeneity makes it difficult to draw definitive conclusions and compare results across studies (Marson & Baldwin, 2019).
Furthermore, some studies have reported contradictory findings. For instance, while Woolery-Lloyd et al. (2010) found that a 5% SAP lotion significantly reduced acne severity, Kircik (2011) found no significant improvement when combining vitamin C with salicylic acid. These discrepancies highlight the need for more standardized research protocols and larger, well-controlled trials to establish the efficacy of vitamin C in acne treatment.
Practical Applications and Recommendations
Based on the available evidence, incorporating vitamin C into skincare routines may be beneficial for individuals with acne-prone skin. However, it is essential to choose stable, high-quality products with optimal concentrations of vitamin C. Pinnell et al. (2001) recommend a concentration of 10-20% ascorbic acid for maximum efficacy, but lower concentrations may be suitable for individuals with sensitive skin.
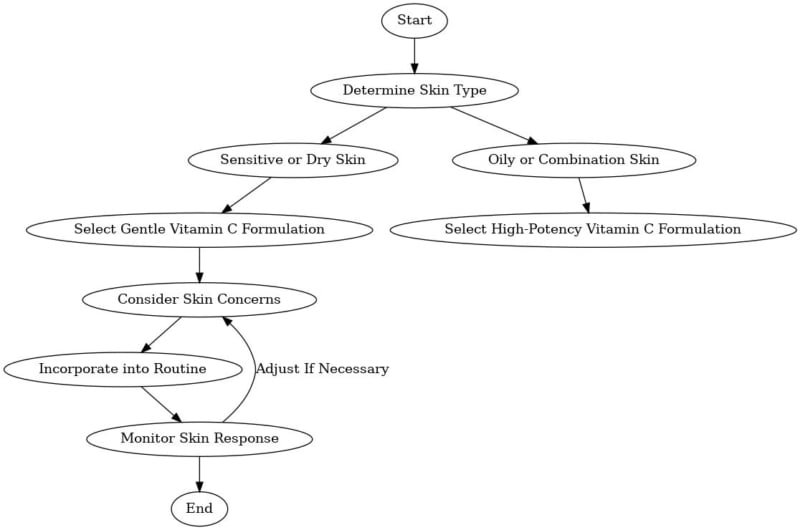
Choosing the Right Vitamin C Acne Product for Your Skin Type
When selecting a vitamin C product, it is important to consider the formulation and packaging. Look for products that are packaged in opaque, airtight containers to prevent oxidation and degradation of the active ingredients. Additionally, storing vitamin C products in a cool, dark place can help extend their shelf life (Al-Niaimi & Chiang, 2017).
Individuals with acne-prone skin may benefit from using vitamin C in combination with other acne-fighting ingredients, such as retinoids or salicylic acid. However, it is essential to introduce new products gradually and monitor for potential side effects, such as irritation or dryness. If any adverse reactions occur, discontinue use and consult with a dermatologist.
Finally, it is important to remember that while vitamin C can be a valuable addition to acne treatment regimens, it should not be considered a sole treatment option. Acne is a multifactorial condition that often requires a comprehensive approach, including proper cleansing, moisturizing, and sun protection. Consulting with a dermatologist can help develop a personalized treatment plan that addresses individual needs and concerns.
Current research suggests that vitamin C can be a beneficial complementary treatment for acne and a valuable addition to skincare routines for promoting overall skin health. Its antioxidant, anti-inflammatory, and skin-brightening properties make it a promising candidate for managing acne and its associated concerns. However, more standardized research is needed to establish the optimal formulation, concentration, and combination of ingredients for maximum efficacy.
As the understanding of vitamin C’s role in acne treatment continues to evolve, future research should focus on conducting large-scale, randomized controlled trials with standardized protocols and outcome measures. Additionally, exploring the long-term effects of vitamin C on skin health and acne prevention could provide valuable insights for both dermatologists and consumers.
By staying informed about the latest research and working closely with skincare professionals, individuals with acne-prone skin can make informed decisions about incorporating vitamin C into their skincare routines for clearer, healthier-looking skin.
CONCLUSIONThis comprehensive meta-analysis highlights the potential of vitamin C as a complementary treatment for acne and its role in promoting overall skin health. The current research demonstrates that vitamin C, with its potent antioxidant, anti-inflammatory, and skin-brightening properties, can help address various aspects of acne-prone skin, including reducing inflammation, sebum production, and post-inflammatory hyperpigmentation.
The studies reviewed in this meta-analysis suggest that different forms of vitamin C, such as ascorbic acid, sodium ascorbyl phosphate, and magnesium ascorbyl phosphate, can be effective in treating acne when used in appropriate concentrations and formulations. Additionally, combining vitamin C with other antioxidants or acne treatments, such as retinoids or salicylic acid, may enhance its efficacy in some cases.
However, it is important to acknowledge the limitations and contradictions present in the current body of research. The heterogeneity among studies, including variations in vitamin C formulations, concentrations, treatment durations, and outcome measures, makes it challenging to draw definitive conclusions and compare results across studies. Furthermore, some studies have reported conflicting findings, emphasizing the need for more standardized research protocols and larger, well-controlled trials to establish the efficacy of vitamin C in acne treatment.
Despite these limitations, the available evidence supports the incorporation of vitamin C into skincare routines for individuals with acne-prone skin. Practical recommendations for selecting stable, high-quality products with optimal concentrations of vitamin C have been provided, along with guidance on storage and potential side effects. However, it is crucial to recognize that vitamin C should be used as part of a comprehensive skincare regimen tailored to individual needs and concerns, rather than as a sole treatment option.
The findings of this meta-analysis have implications for both dermatologists and individuals seeking to manage acne-prone skin. By understanding the potential benefits and limitations of vitamin C in acne treatment, healthcare professionals can make informed recommendations to their patients and develop personalized treatment plans that incorporate this powerful antioxidant. Moreover, individuals with acne-prone skin can use this knowledge to make educated decisions about incorporating vitamin C into their skincare routines and collaborate with their dermatologists to achieve the best possible results.
In conclusion, while vitamin C shows promise as a complementary treatment for acne and a valuable addition to skincare routines for promoting overall skin health, further research is necessary to fully understand its potential and optimize its use. Future studies should focus on conducting large-scale, randomized controlled trials with standardized protocols and outcome measures to better elucidate the role of vitamin C in acne treatment and prevention. Additionally, exploring the long-term effects of vitamin C on skin health and acne prevention could provide valuable insights for both dermatologists and consumers. As the understanding of vitamin C’s role in acne management continues to evolve, individuals with acne-prone skin can work closely with their healthcare providers to incorporate this powerful antioxidant into their skincare routines for clearer, healthier-looking skin.